![]()
Contents>> Vol. 8, No. 1
History that Slithers: Kra-Dai and the Pythonidae*
James R. Chamberlain**
* I would like to sincerely thank Professors David Holm, Meng Yuanyao, and Weera Ostapirat for their kind assistance in the preparation of this paper.
** Center for Research on Lao Languages, National University of Laos, Dong Dok, Vientiane, Lao PDR
e-mail: jimchamberlain[at]gmail.com
DOI: 10.20495/seas.8.1_25
This paper brings together a number of disciplines in order to demonstrate how historical, anthropological, ecological, zoogeographical, ethnobiological, and linguistic evidence relating to the physical distribution and linguistic representations of pythons in northern Southeast Asia and southern China can be brought to bear on Kra-Dai prehistory and intrafamilial as well as interethnic relationships. The normal and most recognized word for ‘python’ is confined to the Tai family proper, and even then there are some qualifications. Two species of python are found in much of the Tai linguistic area south of the Sino-Vietnamese border, but only one, the Burmese python, occurs in Guangxi, Guangdong, and Hainan. Some Central Tai dialects have acquired another name that seems to be Austroasiatic (AA) in origin, and yet no AA languages are found in those areas. It is suggested that these dialects received the word via Kra to the west. On the eastern side, yet another surprising correspondence is noted between Lung Ming in southern Guangxi and Hlai on Hainan. Sek, located far to the south, which usually preserves archaic forms of Be-Tai, has no words for ‘python’ that correspond to those in the rest of the family. Close examination of the linguistics of this particular member of the Southeast Asian mega-fauna reveals a pattern of interaction between the families of the Kra-Dai stock, Austroasiatic, and southern Chinese that mirrors the phylogenetic tree.
Keywords: Kra-Dai, Tai, ethnozoology, python, Tai history, Chinese history, Vietnamese history, Southeast Asian history
I Introduction
This study discusses pythons in southern China and northern Southeast Asia, the interesting historical linguistic situation that surrounds them, and the implications of this for reconstructing certain aspects of the Kra-Dai past. Using linguistic evidence as the starting point, comparative evidence is offered from a number of fields such as zoogeography, ethnobiology, history, folklore, and anthropology, providing an example of how bringing all of these fields of study together can provide insights into patterns of settlement and distribution of families within the Kra-Dai stock.
Specifically, it will be argued that,
1) The Proto-Tai (PT) etymon for ‘python’ *hnɯam A referred to the Burmese python (Python bivitttatus).
2) The Siamese forms distinguishing two species of pythons are an independent innovation, related to the sympatric distribution that is particularly evident in central Thailand. The only other language where /laam/ is found is Yuan (in northern Thailand) where it is the only taxon for python.
3) Old Chinese borrowed the Proto-Tai (PT) form from the Eastern Ou Yue people in Zhejiang.
4) Cantonese and perhaps other southern Chinese contact words for python were borrowed at a later date directly from Northern Branch Tais in Guangdong and Guangxi.
5) The Central Tai (CT) taxon Proto-CT *daaŋ B derives ultimately from Austroasiatic (AA) via Kra.
6) There exists a Tai-python < AA-skink association that may be seen in discontiguous localities, in both CT and Sek, this seems to have been an avoidance mechanism in areas where pythons were associated with taboos.
7) The Lung Ming-Hlai correspondence, Lung Ming /laan C4/ ~ Proto-Hlai *C-naaɲʔ, may be an original Daic form that was borrowed into Old Chinese *mâŋʔ < *malâŋʔ (Schuessler 2007).
8) Chinese medical uses for pythons developed independently and were not borrowed from Tai peoples.
9) Ou-Yue Tai (CT and Northern Tai [NT]) beliefs associated with pythons are spiritual in nature, whereas Lo-Yue (Southwestern Tai [SWT]) associations are primarily related to edibility. This can be seen in the naming practices evident in the folk systematics.
10) A term for python cannot be reconstructed in Proto-Kra-Dai, and as a result we should look for a python-free homeland for this stock.
Language is prior. For purposes of this paper, language sets the backdrop against which other types of information may be placed. The methods of comparative and historical linguistics are based upon immutable laws such as the regularity of sound change leading to reconstructions of proto languages that date back to prehistoric periods. There are no parallels in the fields of history, anthropology, psychology, and so on. Historical linguistic studies lead to the establishment of classifications that show relationships between the branches and subgroups of linguistic families and stocks, and these in turn may be linked to historical or archeological records, depending upon extant evidence. The comparative method begins with the assembling of data from living languages or in some cases from written records, and the identification of cognates, morphemes that share a common root, in order to examine the variation in consonants, vowels, tones, and meaning that allows the linguist to reconstruct the original form. Take the words for ‘moon, bile, flower’, Siamese /dɯan, dii, dɔɔk/ but Black Tai (spoken in northwestern Vietnam) /bɯan, bii, bɔɔk/. The correspondence of d– and b– poses the problem of reconstruction of the proto initial consonant and has led linguists (such as Li 1977) to suggest, based on the common point of articulation of d and l, an original *bl– cluster for these items and other words with similar correspondences. Later, with the discovery of Sek, a language that had remained isolated in a remote part of Khammouane Province in Laos, the reconstruction was confirmed in the forms /blian, blii, and blɔɔk/ that has preserved the original clusters (Gedney 1970, 83). This is a simple example but serves to demonstrate the cogency of the method in terms of predictability and in providing a foundation for the present study.
Zoogeographic areas here concern the Oriental Region of the Paleotropic Realm, bounded to the north of the Yangtze by the Palearctic Region of the Holarctic Realm, and to the south by the Australian on the southern and southeastern side of the Wallace Line, so-named by Thomas Henry Huxley, that follows the deep water channel between the Sunda region (Java, Bali, Sumatra, Borneo, and Mainland Southeast Asia) and the Sahul (Australia and New Guinea) continental shelves separating Oriental fauna such as tigers and elephants from the Australasian marsupials (Darlington 1957; J. McKinnon and K. McKinnon 1974; Müller 1974). The transitional area between the Wallace and the Zydekker Lines is known as Wallacea and there is a small zone here where the two pythons, bivittatus and reticulatus overlap. Otherwise bivittatus is not found this far south. To the north in China, at least as far as Anyang in Henan Province, Shang Dynasty remains of elephants, rhinoceros, and tapirs have been found, indicating that original sub-tropical forests in this area were apparently destroyed by extensive human settlement (Müller 1974; Chi 1957). But to my knowledge so far, python remains have not been found at this or other sites this far north. By comparing the distribution of animals with linguistic representations and cognates across language families, hypotheses can be formed with respect to original homelands and migrations.
Ethnozoology, the study of human-animal relationships, is the branch of cognitive anthropology that, among other things, seeks to uncover ethnocentric epistemologies underlying indigenous classifications of animals. Here we add to this a diachronic dimension so that what may be compared historically are sets of classifications which may or may not agree across families and branches within Kra-Dai. Folklore fits here as well, and in principle, such anthropological questions as gender and kinship aspects of animal naming and the degree to which this corresponds across languages and families. The rather positivistic approach of the folk biological taxonomists such as Berlin (1972; 1992) utilizes a system of terminology analogous to the Linnaean system, based entirely upon physical characteristics. The terminology is useful but does not easily accommodate the non-physical dimensions of taxonomies without including additional information (Chamberlain 1977; 1992; Aung Si 2016).
History begins with writing and the preservation and availability of written sources. For the earlier periods these may or may not be associated with identifiable ethnicities currently recognized. In our focal area of southern China, we are fortunate in that numerous ancient texts have been preserved and are available for interpretation by historians.
II The Pythonidae in Northern Southeast Asia and Southern China
Pythons are nonvenomous old-world constrictors inhabiting Africa, Asia, and Australia. Members of the family Boidae are also constrictors but are found mostly in the new world, primarily in Central and South America and the pacific islands. (There are some exceptions to this generalization, a python species found in southern Mexico and Costa Rica and a boa found in West Africa [Encyclopedia Britannica].) Pythons are oviparous, but the Burmese python may sometimes reproduce by parthenogenesis (Groot et al. 2003), the only snake of this family known to do so. Primarily nocturnal, pythons feed on mammals and birds and occasionally on reptiles.
The family Pythonidae in northern Southeast Asia is represented by two distinct species (see Fig. 1), the Burmese python, Python bivittatus, and the reticulated python, Malayopython reticulatus (formerly genus Python). A third species, Python molurus, the Indian Python, confined to South Asia, closely resembles bivittatus, which for many years was considered a subspecies of molurus. The reticulated and the Burmese pythons are two of the three largest snakes in the world, in terms of both weight and length. Each may weigh over 150 kilograms and measure in excess of six meters in length. One reticulatus specimen is reported to have attained a length of more than nine meters.

Fig. 1 Burmese Python (Left) and Reticultated Python (Right)
Source: Wikipedia Commons.
As can be seen on the maps, the Burmese python has a decidedly more northerly distribution than that of reticulatus, on the east reaching as far as Guangxi, Guangdong, Fujian, and possibly eastern Jiangxi. On the west the range stretches to central Yunnan and even to southeastern Sichuan. On the mainland the southern boundary is Surat Thani, Thailand, but in the islands, it occurs in Java, Bali, and southern Sulawesi.
The northern range of the reticulated python stops approximately at the Chinese border with Vietnam, Laos, and Burma, and extends westward across all of India. It is found throughout insular Southeast Asia. It can also be seen on the maps that where the islands are concerned, the two species are mostly in complementary distribution, with bivittatus being found on Hainan, but absent from Malaysia, Indonesia, and the Philippines, the only exception being Java and southern Sulawesi where the two species again overlap. Neither species is found on Taiwan, though the Burmese python is found on the Kinmen Islands.
Burmese and reticulated pythons are sympatric throughout much of mainland Southeast Asia south of the Chinese border (see Fig. 2). But ecological niche partitioning is difficult to define. Bryan Stuart (personal communication [p.c.]) notes that reticulated pythons are usually associated with water sources, while Burmese pythons are sometimes not. In Indonesia bivittatus is said to prefer more arid locations and is less common near human habitations (Stuart et al. 2012). In addition, bivittatus can tolerate cooler climates and is known to brumarte (hibernate). Otherwise description of their habitats is more or less the same, especially as to their being found in and around water sources, streams, and ponds. Burmese pythons are common around the Tonle Sap in Cambodia, and in the Erhai Lake in central Yunnan. But on the island of Bali in Indonesia, Burmese pythons are found mainly in the northern, more arid part of the island, whereas reticulatus is restricted to the more humid areas of the center and south (Mark Aulia p.c.). The Burmese python population throughout its native range is in serious decline (Stuart et al. 2012), even while it has increased dramatically in the Florida Everglades where it is thriving as an introduced species following Hurricane Andrew which destroyed a nearby breeding facility in 1992 allowing the escape of the captive snakes.
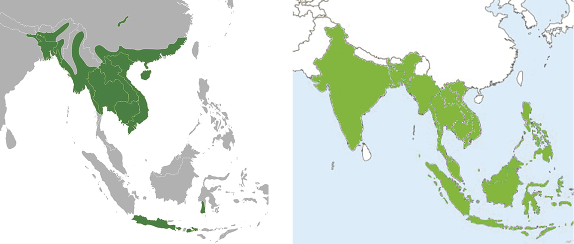
Fig. 2 Ranges of Bivittatus (Left) and Reticulatus (Right)
Source: Wikipedia Commons.
In Siamese, the reticulated python is called /ŋuu A4 lɯam A1/ งูเหลือม ‘snake + python’ and the Burmese python is /ŋuu A4 laam A1/ งูหลาม, distinguished as two separate species in the lexicon but obviously deriving from the same root.
In Indonesia, Malaysia, and the far south of Thailand, there exists yet another species known as the blood python (because of its color) Python curtus. It is much shorter and heavy-bodied, with a stubby tail, but because of its southerly range it is not relevant to this paper, except in one respect: its Thai name งูหลามปากเป็ด ŋuu laam paak pet or ‘duck-billed python’ uses the bivittatus taxon, even though the Burmese python does not occur in southern Thailand where curtus is found, an indication that the two separate linguistic variants must have developed prior to the time that Thays moved into the peninsula (see Fig. 6 below). This situation and its implications will be discussed in greater detail below.
That the names for python in Kam and Sui languages are not cognate with each other, nor with the Tai forms is interesting. Unfortunately, our data from these areas is so far incomplete.
Python bile was the quintessential medicine of southern China and trade in the reptiles became a lucrative enterprise for local people. Schafer (1967, 217) notes that at least as far back as the Tang Dynasty, python farming was carried out by specialists in the counties of Lo and Lei (in present-day southern Guangdong). These were no doubt Burmese pythons that inhabited this region.
With the exception of Siamese not many languages have differentiated them. It may be true, however, that for the second python etymon found in Central branch dialects, for example Lung Ming /taaŋ B/, that is, in the same area, that this is a possibility. One such example from Tày Nùng in Vietnam was brought to my attention by David Holm and is discussed below.
III Kra-Dai (Kra-Tai) Languages
The languages of this stock include three main branches and two sub-branches as shown in Fig. 3.

Fig. 3 The Kra-Dai Linguistic Stock
More recently Weera Ostapirat (2018), as shown in Fig. 4, has modified this arrangement slightly to show the closer affinity of Hlai to Kam-Tai, positing an initial bifurcation between Kra on the left and Daic on the right, and a secondary split between Kam-Tai and Hlai. He also shows Be as splitting off at an earlier period.
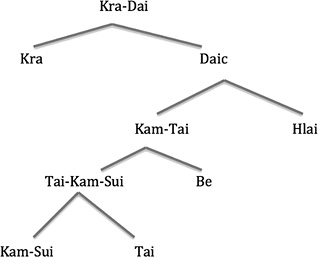
Fig. 4 Modified Kra-Dai Tree
The present-day locations of these main groups can be seen on the map in Fig. 5.
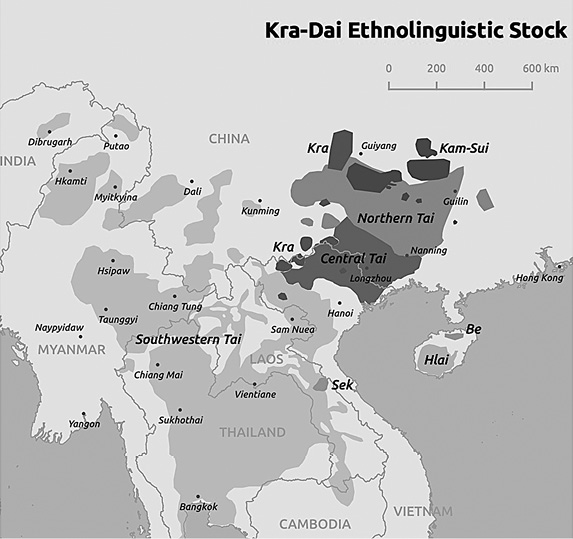
Fig. 5 Map of the Main Kra-Dai Ethnolinguistic Groups
Source: David Wharton, James Chamberlain, and Sylvain Dorey, public domain.
It is possible, based on linguistic evidence, to suggest routes of migration that coincide with history to the degree that written records are available. For Chinese this usually means beginning in the Han Dynasty (201–220 BCE), but for Kra-Dai it has meant that written languages are located mainly to the west, beginning in the twelfth century CE.
Work by David Holm (e.g. 2015) with written Tai vernaculars using Chinese characters is most promising and pushes back the time depth for Tai literacy in Guangxi and adjacent parts of Vietnam. In fact, Holm (2017) dates the early vernacular scripts of Central Guangxi to the time of the Sui-Tang transition (c. 600 CE) but with even earlier strata dating from the Han, and suggests that the earliest strata of Chữ NômTày may date from this same period, that is six or seven centuries before the appearance of Vietnamese Chữ Nôm. Still the bulk of Tai history is focused on the western and more recently settled Southwestern Branch groups, skewing the big picture and misdirecting attention away from the geographically older and historically more relevant territory of Lingnan and the southeast China coast.
An example of how the phylogenetic tree relates to history of the Tais can be seen in Fig. 6.
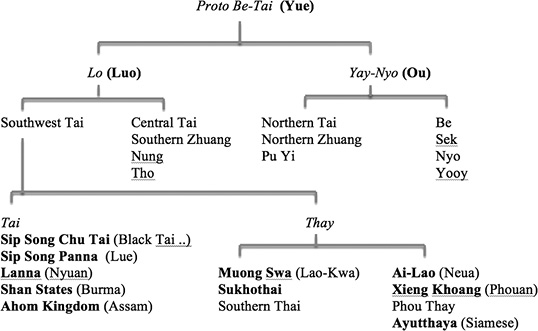
Fig. 6 The Tai Family Tree Linked to Historical Tai States
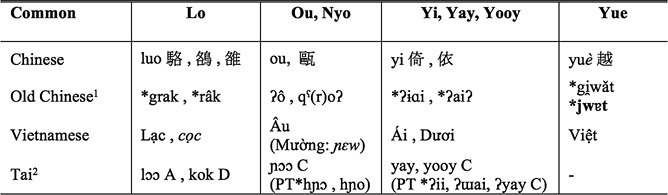
Note here that in the upper branchings, ethnonyms from Chinese history can be related to autonyms of modern Tai groups. The above Table 1 may assist readers in following the often-confusing array of ethnonyms as used variously in the historical sources.
Table 1 Tai Ethnonyms in Chinese and Vietnamese Histories
IV Snakes and Pythons in Ancient China
Eberhard (1968, 150) suggested that “among the tribes in Southeast Asia, where Ku-like magic is very widely known, the snake may be the true and only magic power.” Thus the category of snakes in Kra-Dai, as might be expected, is a complex one.
Some snakes are named with disyllabic generic level lexemes (like lizards) which, as I have suggested elsewhere (Chamberlain 1981), were probably associated with ku [pinyin gu] magic. Snakes associated with ku magic were mainly kraits and pit vipers which have disyllabic names. Eberhard (1968, 159) writes that on the fifth day of the fifth moon the five dangerous animals—toad, snake, gekko, centipede, and scorpion (or spider)—were placed in a pot to fight with each other, the victorious one becoming the most powerful for that year and the one used for ku poison and magic. I have discussed the matter at some length in Chamberlain (1993) suggesting that the Tai etymon for ‘millipede’ /(boŋ C3) kɯɯ A/ [/boŋ/ ‘caterpillar’] is a possible correspondence, found throughout SWT and CT. Millipedes are similar in appearance to centipedes which are said to be enemies of snakes and an antidote for ku poison.
Also, according to Eberhard (1968, 380ff), snake cults flourished along the coastal areas of southern China in Zhejiang, Fujian, and Guangdong, in other words in the northerly range of the Burmese python. Eberhard also mentions (ibid., 381) that in Guangdong and Guangxi the word ‘snake’ itself was taboo, though unfortunately we do not know what that word was. Snakes could marry women and be worshiped as river gods or gods of lotus ponds. And it is probably not accidental that the Burmese python is usually found in or near water sources. Snakes in such areas are also associated with good health, good fortune, and wealth. All of these beliefs and more are associated by Eberhard with his “Yueh” Culture, along with bronze drums and the dragon boat festival, linking Tai and Yue (see Chamberlain 2016).
Many of the Tai and other Kra-Dai beliefs concerning pythons are very similar to one another. For example the Black Tai say that if the vine called /cɯaʔ num nɛɛ/ (botanical identity not known) is used, the python can be led about like a dog on a leash, not unlike what Swinehoe wrote in Hainan (1870, 241) that, “the natives declare that they are not hurtful to man, and are easily caught by throwing over their heads a noose of twisted grass, and with this may be led about without danger.”
In the eastern range of Tai, in northern Vietnam and China, pythons are not normally classed as snakes and are referred to as simply *tua A (animal) + ‘python’. The term ‘Snake’, as discussed at the outset, belongs to that category of organisms called Life Forms (LF) by the folk biological taxonomists, along with ‘fish’ and ‘bird’. It has been suggested (Chamberlain 1977) that what sets these categories apart is their lack of physical resemblance to human form, such as the lack of arms and hands, or bodies that are covered with feathers or scales. Where members of such categories as defined by form become closer to humans due to physical or psychological proximity they lose their classificatory status as bird, fish, or snake. The chicken, the duck, and the crow cease to be birds, and the python is no longer a snake.
Although it is convenient here to use the folk systematics terminology for purposes of clarity, this does not imply that school of thought has been accepted verbatim. For example, the use of LF to include or exclude certain species is not well developed, and some LF categories that are put forth in the literature are not in fact used in real language. An example would be ‘mammal’ in English where only recent technical scientific words are available. We do not say, *squirrel mammal, or *wolf mammal in our speech, whereas we do say ‘bull snake’ or ‘rattlesnake’, employing the LF as a part of the taxon. Yet the orthodoxy would assign the label LF to both mammal and snake.
In addition, animals classified by LF, may be less subject to rituals and sacrifices or other edibility restrictions. For example in Lao, presence of a LF in the name may mark the difference between domestic and wild, as in
/too pet/ ‘UB (animal) + duck (G) = domestic duck’
/nok pet/ ‘LF (bird) + duck (G) = wild duck’
Or, also in Lao, the traditionally inedible soft-shell turtle is reclassified as a fish in order to justify its edibility. Thus,
Black Tai /too faa/ ‘UB (animal) + soft shell turtle (G)’
Lao /paa faa/ ‘LF (fish) + soft shell turtle (G)’
In most Central (CT) and Northern branch (NT) languages, pythons are not classed as snakes, that is, whereas in Southwestern Tai (SWT) the name is a combination of the LF marker ‘snake’ and the Generic (G) level word ‘python’, in the CT and NT languages the structure of the taxon is Unique Beginner or Kingdom (UB) plus G, that is, ‘animal + python’. (For additional details see Berlin 1992.) A similar problem is addressed by Bulmer (1967) in his classic paper, “Why Is the Cassowary Not a Bird?” and no doubt as in that case, the answer is not straightforward where pythons are concerned. Bulmer concluded (1967) that the cassowary’s classificatory position cannot be attributed to taxonomic, morphological, or behavioral characters alone, but must be seen in terms of the culture or cosmology of the Karam ethnic group as a whole. For the python and the Tais (and perhaps other families of the Kra-Dai stock) the great size and its association with water and women, sets it apart from other snakes and other animals generally. We have no firm grasp of either synchronic or diachronic ethnographic detail related to pythons among Tai groups, only the smatterings of information to be gleaned from random fieldwork and Chinese historical documents. The anomalous position it came to hold, neither fish nor fowl to use the English expression, due both to its mythological role as deity, brought it firmly into the human domain. In some areas its name was elevated to become the LF marker for ‘snake’, a quintessential semantic shift (David Holm p.c.).
Of great interest is Eberhard’s association of the snake (and in most cases I believe we can relate his ‘snake’ with the python) with the “Yüeh” culture, as was the case in other early sources discussed below. I will not go into the problems issuing from his cultural divisions, but his summation is interesting when we think of the probable identification of Tai with Yue on China’s southern coast, the home of the Burmese python. He relates that for his Pa and Yao cultures, the snake is inauspicious or evil. In his Thai culture, snakes are equated with the lung-type dragon and with sexual symbols, but, “only in the Yüeh culture is the snake considered a veritable deity, mostly benevolent. It can marry humans but also receive humans as sacrifices which are rationalized as symbolic marriages. In his Yüeh culture, the snake is most importantly a river god, closely related and often identical with the water dragon chiao” (Eberhard 1968, 386), an identification usually reserved for the salt-water crocodile, Crocodylus porosus, PT *ŋɯak which in most Tai languages has become a mythological creature (see Chamberlain 1977, 67ff). This crocodile has related forms borrowed into Chinese along the coast, such as Cantonese ngok, or Amoy go’k-hî (Douglas 1899). (Benedict in 1976, to account for the dipthong in Tai, proposed *ŋwɯak < ŋ(i)(w)ak based on Indonesian *maŋiwak ‘shark’.) And, supporting Eberhard’s linking of ‘dragon’ with serpent, Lung Ming, a CT language, has /tuu A2 ŋoo A4 yii A4/ for ‘crocodile’, that is, classed as a snake (ŋoo).
Snakes, as they are the world over, are associated with various other beliefs and Southeast Asia is no exception. There is the snake that eats the gekko’s liver, the snake that leads the buffalo away, the snake that strikes with its rear end, the snake that throws clubs in the forest. The most universally feared snake (though completely harmless) is Typhlops, the blind snake that lives underground (Chamberlain 1981).
Following the Chinese colonization of the south beginning in the Qin and Han dynasties, pythons gradually became recognized as the source of valuable medicine made from their fat pads and bile, and thus became economically important, ostensibly a contribution of the southern peoples to the medical repertoire of Chinese physicians (Churchman 2016, 157ff). The Tai word for ‘fat, healthy’ PT *bɯːj or *buːj A (Ostapirat 2013, 197 Table 16) may be a loan from OCM *bəi 肥, perhaps as a result of its association of python fat in Chinese medicine. But, given the python’s southern distribution, one would assume that Chinese medicine most probably acquired the knowledge from Tais, in which case the borrowing may have gone in the other direction. The Black Tai of northwestern Vietnam, for example say that the fat /pii A4/ of the python /too A2 lɯam A1/ can be used as a substitute for castration when smeared on the penis and testicles of a horse. They also say the the gall bladder /bii A3/ of the python is placed in alcohol and taken to cure pain in the bones (Author’s fieldnotes 1977). I do not know whether any such specific uses are mentioned in the Chinese materia medica.
Nevertheless, while the idea that the Chinese received their python medical knowledge from the Tais is appealing, there is a complication in this interpretation. David Holm (p.c.) reports that while the Chinese materia medica and pharmacopoeias are replete with references to pythons, the local Tai or Zhuang texts contain no mention of medicinal uses of python parts. For example, medical uses of bivittatus and some 15 other snake species are documented in Lin Luhe 林呂何 (1991, 310–348), a Chinese text. However, there is no mention of the python in the Zhuang materia medica (Huang Hanru 2001; Liang and Zhong 2005; Wang Bocan 2006; Zhu 2003), nor in the medical manuals of the Bouyei (Pan et al. 2003), Sui (Wang Hou’an 1997), Gelao (Zhao et al. 2003), and Yao (Qin and Li 2001). “The Guangxi medicinal animals book is evidently about Chinese (Han Chinese) medicine, and contains no references to minority medical systems” (Holm p.c.).
Fan Chengda (1984), writing in the Song Dynasty, relates that, “The southern people hunt them for their skins, scrape off the scales, and stretch them over drums” (translation by David Holm p.c.). From this it may be inferred that while Chinese interest in pythons was restricted to practical medicinal value, Tai interest lay mainly in the value of these snakes in ritual (as in the sacred importance of the sound of drums), likely related to Eberhard’s depiction mentioned above of snakes in his Yueh culture as the subject of myth and veneration. Thus a neat parallel may be seen in the medical importance of the python to the Chinese, in juxtaposition to its spiritual importance to the Tais.
It must be the case, however, since we can assign the gloss of ‘Python bivittatus’ to PT *hnɯam, that Chinese, a northern language, did not have a word for python until their contact, in whatever form, with Proto-Tais (see below).
The flesh of pythons was also consumed, but only selectively. Edibility of pythons and other snakes is not universal. In Western Nung snakes classified as /nguu A4/ cannot be eaten; only the python, not classified as a snake, is deemed edible. For the Yay, no snakes are eaten as they are considered bad omens and if a snake enters the house it will cause illness. The same is true for Black Tai with respect to kraits or king cobras if they are seen during the day (Chamberlain 1981).
V Linguistic Forms for ‘Python’ in Kra-Dai and Neighboring Languages
By far the best-known terms for ‘python’ in Tai languages are reflexes of PT *hnɯam A. In general, for the Southwestern branch the initial is always l- and in the Central, Northern branches it is n-. Borrowing into southern Chinese dialects has been assumed by some, for instance Churchman (2016, 158). But Pulleyblank (1991, 264) reconstructs Early Middle Chinese (sixth century) EMC *ɲiam [A] (the pinyin rendering is rán) with a voiced initial, indicating the borrowing must have taken place earlier. For Cantonese, 蝻蛇 naam4 [=Tai C4 tone] se4 ‘python + snake’ is the normal word, also in the voiced series, the C tone suggesting Cantonese may have borrowed directly from Tai, based on pitch contour similarities at the time, as opposed to being a regular development from EMC.
The forms in Be, such as Zhang’s /num55 A2/, and Savina’s niem, also suggest early Be voiced initials *n- or *ɲ– (Ostapirat p.c.), rather than *hn- or *hɲ– inherited from a hypothetical Be-Tai. The form would therefore seem to derive from a pre-Proto-Tai source.
Furthermore, there is indeed confirmation that borrowing into Chinese must have occurred at a time earlier than EMC. For example, David Holm (p.c.) notes that the 100 CE Shuowen jiezi entry for python reads:
蚦,大蛇,可食。从虫冄聲。人占切。
Python, a large snake, edible. [Graphic composition] follows ‘insect (虫), with ran (冄) as a phonetic element. [Pronounced with the initial of] ren (人) cut with [the final of] zhan (占). (Shuowen jiezi 13A: 16a, 1963, 278)
(Ran, ren, and zhan are of course modern Pinyin forms, not reconstructed Old Chinese, and these last three characters of the entry, indicating the pronunciation, were apparently added at a later date.)
An even earlier occurrence of the ran character is found in the Huainanzi 淮南子 (c. 139 BCE) where it is written, “When the people of Yue get a python they regard it as an item of highest value” (fascicle 18 Jingshen xun 精神訓). Here the character used for python is 髥, yet another variant with the same pronunciation.
These would seem to be the source for Zheng Zhang’s OC *njam 蚺 ‘python’ (2013, 450). Holm cautions that the character listed in the Shuowen jiezi is 蚦, not 蚺. The second is a later variant graph first used in the Three Kingdoms period.
Passages from the various post second–third century CE recensions of the Yi Zhou Shu 逸周, the original early Zhou text which is no longer extant, are worth examining for their associations of snakes with the Eastern Ou or Ou Yue (Huang Huaixin et al. 2007). The chapter entitled “Wang hui jie” 王會解 “The Royal Audience Explained,” fascicle 7, describing the Ou people.
The people of Ou, cicada snakes. Cicada snakes are tame, and delicious to eat.
歐人蟬蛇。蟬蛇順,食之美。(834)
On this passage, the earliest commentator, Kong Chao 孔晁 of the Wei-Jin period (fourth–fifth century CE), identifies the Ou as a particularly ophiophagous group: “These are the Eastern Yue Ou people. Their snakes are especially numerous even compared with Jiaozhou, and are a high-class delicacy.” The Eastern Ou were located in the Ou River valley near Wenzhou in present-day Zhejiang. They are furthermore considered by some to belong to the Tai linguistic family (see Chamberlain 2016).
Another commentator Pan Zhen 潘振 explains: “The cicada snake is the python. Its tail is round and without scales, and on its body it has spots and stripes, like old dark knotted embroidery” (835). Note that Eberhard (1968, 187), though he provides no details, mentions a “cicada-god” (ch’iung-ch’an) associated with the south.
It is still puzzling nonetheless because the Huainanzi and the Shuowen jiezi are northern Chinese works, that is, they were compiled north of the Yangtze well beyond the range of the Burmese python. One explanation is suggested by David Holm (p.c.), that pythons, as well as other exotic animals and oddities were sent to royal courts in the north as tribute gifts and that the names of the animals as well as the names of the people were recorded at these times. Descriptions of such presentations were described in the Yi Zhou Shu mentioned above and certainly seem plausible despite problems with the dating. Most importantly, neither the Huainanzi nor the Shuowen jiezi, nor the Yi Zhou Shu recensions make any mention of medicinal use of the python by southern (non-Chinese) peoples.
For Proto-Tai Li (1977) reconstructs PT *hlɯem noting the l- ~ n- variation is sporadic and rare in the voiceless series, though in fact the variation is essentially SWT *hl- , CT and NT *hn-, perhaps implying an initial cluster or a pre-syllable, but more likely a SWT innovation as opposed to an indication of variation at the PT level as in Li. This would imply PT *hnɯam ‘Python bivittatus’ (Ostapirat p.c.).
Vocalic reflexes are:
| SW | -ɯa- ~ ɤɤ- ~ -aa- |
| C | -u(u)- ~ -ua- |
| N | -ua- ~ -ɯɯ- ~ -uu- |
| Be | -ie- ~ -u- |
With respect to the vowel, Ostapirat (2013, 203) reconstructs a short dipthong *ɯa based on the correspondence of Siamese –ɯa– and Po-ai –ɯɯ– in such words as
| Siamese | Po-ai | |
| moon | dɯan | nɯɯn |
| blood | lɯat | lɯɯt |
| crocodile | ŋɯak | ŋɯɯk |
But he also (ibid., 223) notes the possibility of a long labialized dipthong *wɯaː giving:
| Siamese | Yay | |
| boat | rɯa | rua |
| salt | klɯa | kua |
In some C and N dialects that show rounded vowels, Ostapirat (p.c) suggests this could be the result of assimilation of the vowel to the point of articulation of the final labial -m, as for example, *ɯa > ua (> u:/u). He notes a similar correspondence in a separate etymon, *hlɯam C ‘brilliant, shiny’ where the Northern dialect Yay has /luam/, as another example.
The occurrence of /laam A1/ in Siamese and Yuan (Sanguan 1969) remains problematical, but could very well be simply a variant that appeared when the early SW Tais came into the territory that is now Thailand where both species were abundant. Given its more northern distribution, we assume that PT *hnɯam must have referred to Python bivittatus. At some point in what is present-day Thailand, the referents of two linguistic forms, /laam/ and /lɯam/, must have become reversed, though not before the Thays (before the establishment of Siam as the nation-state with a Thai autonym) had moved south and applied the /laam A1/ taxon to Python curtus, the blood python. As reticulatus is found more frequently in and around human habitations, it probably became the default species, replacing bivittatus. The Yuan form mentioned for northern Thailand, preserves only the /laam/ variant as the sole word for ‘python’.
Such variation in the same language from a single etymon is reminiscent of the situation in Laos, in the Xam Neua and Pak Xeng dialects of Thay Neua /mii A1/ and /mɯay A1/ which have come to represent different species of bears, Ursus thibetanus and Helarctos malayanus (Chamberlain 1977, 126).
Lung Ming, as shall be seen, is another Tai language that retains two distinct etyma for ‘python’ but neither form is a reflex of PT *hnɯam A.
In this regard a second form needs to be considered, reflexes of Proto-Central Tai *daaŋ B, found in Lung Ming, Lei Ping, Lung Chow, and I suspect other locations in southern Guangxi. Notably the words for various species of lizards in Austroasiatic languages contain the form /daaŋ/ or /taaŋ/, for example Khmu Am /daaŋ trpaak/, or Khmu Yuan /tàaŋ lʔiər/ ‘skink’ from an original voiced stop. Savina’s Tay also has gìưng tạng lùm for a ‘small lizard’. However, since there are no AA speakers in the areas where these Tai dialects are spoken, there would need to be an explanation for how this possible contact form emerged. Finally, some Northern and Central Tai languages, classify skinks as snakes (skinks are long shiny lizards with small limbs), for example Yay /ŋɯa4 sii1 keem1/ thereby completing the snake-lizard-*daaŋ linkage but leaving us with a puzzle.
I suggest that the answer lies in Kra as its former territory seems to have covered the areas in question prior to the arrival of the Tais (cf Chamberlain 2016). They are also known to share some lexicon with AA (Ostapirat p.c.), hence the Red Gelao form /ma34 ŋ̍44 ʔɗaŋ44/ for ‘python’ clearly indicates such a connection. It also supports the hypothesis that the Kra spread from west to east. Borrowing from AA could explain the tonal aberrancy as well.
Cantonese tang4 , (Mandarin téng) 螣 ‘flying dragon’ is most likely not the source as it has a short vowel, the wrong tone class, and difficult semantics. In addition the word seems not to have existed in OC.
Only one source differentiates *daaŋ B and *hnɯam A in the same language, and that is the Tày Nùng dictionary of Hoàng Văn Ma et al. (2006). It is not clear, however, to what extent dialect variation may account for this since both nưưm, and lươm are cited in addition to taang indicating considerable dialect differences under the rubric of “Tày Nùng.” And although the glosses for these in Vietnamese are ‘python (con trăn)’ and ‘earth python (trăn đất)’, the species identification is not provided. The situation is worthy of consideration however since according to the range maps both species co-occur in northern Vietnam.
David Holm and Meng Yuanyao (p.c.) note that /taaŋ/ (dangh) in some areas of Guangxi may mean simply ‘snake’. This type of upward semantic shift, what folk biological systematicists would call a diachronic development from Generic to Life Form taxa, is relatively common, and might even have been anticipated, where one conspicuous and culturally important animal or plant becomes the general term. For example, the word for ‘oak’ becomes the word for ‘tree’ in some Native American languages (Berlin 1972).
Located between Hmong-Mien in the north and Austroasiatic in the south, Kra has an old and mostly unrecognized history. Its name is related to the ancient Chinese states of Xia and Chu (cf Chamberlain 2016, 39ff; Miyake 2018, 18.5.11.23: 59).
Xia (*graʔ C), Chu (*tshraʔ C), and Kra (*khra C)
it appears in a number of important historical contexts in etymologies of which most scholars are unaware, for example:
OC *krâuɁ > Jiaozhi > Giao Chi ‘KEO’
*kra C > *krau C ‘GELAO’ or ‘Ch’ü-lao’ 屈撩 [6th c.]
PKD *khra C > PK *kra C > krau > lau ‘KLAO’ > laau ‘LAO’
Another example of a shared word with AA is Kra /klaaŋ/ ‘hawk’, PMK *k(a)laaŋ ‘osprey’.
Lung Ming (Gedney fieldnotes) has yet another word glossed as ‘python’ /ŋow4 laan6 C4/ which is cognate with Hlai /naaɲ ~ naan C/ indicating a contact to the east, in keeping with the assumed closer alignment of Hlai with Kam-Tai. The word is reconstructed in Proto-Hlai as *C-naaɲʔ (=Tai C tone) but so far cognates in other languages have not been found. Hlai (Li) peoples occupied the coastal regions from Canton to at least as far south as Jiuzhen (Thanh Hoa and south) and the Central-Southwestern Tais (the Luo Yue), and the Northern Tais (the Ou Yue) would have encountered them when they migrated into the area fleeing the Qin and Han invaders. It is generally assumed that Hlai is closer to Tai than Tai is to Kra, so these terms may perhaps descend from a Proto-Daic root such as *mlaaɲʔ, to use Ostapirat’s classification. It is tempting to infer that a suggested Proto-Daic form is the source for OC *mâŋʔ < *mlâŋʔ (Schuessler 2007). The tone category matches, and note Amoy: bóng, báng ‘python’ (Douglas 1899). On the Chinese side, I do not know whether reflexes for this form occur in the non-coastal Chinese dialects. It does not appear in the OC reconstructions of Baxter and Sagart (2014).
Sek etyma are of interest as they imply that Sek may have split off from the Tai mainstream earlier than has been assumed. Given that this language preserves so many other archaisms it is puzzling that Sek contains none of the forms mentioned so far. For Sek /ŋua4 tlɔɔ1/ the origin would seem to be Kri-Mol: Kri: /tălɑ̤ɑ̤/ and Mlengbrou: /tăloɔ/ both meaning ‘skink’ (Chamberlain 2018, 109). But note also Mlengbrou /kra̤w/ ‘python’ (ibid., 107). As mentioned above, there seems to be a connection between python and skink in naming practices. The other Sek word /ŋua4 trɛn1/ is an obvious Kri-Mol (Vietic) loanword, for example Ahlao /luk tǎlɛn/ from PMK * t( )lʌn (Diffloth 1980). Sek and Kri-Mol-Mol languages of Kri, Mlengbrou are located on the Nakai Plateau in the Nam Noy and Nam One river valleys of Khammouane province in central Laos.
In Kam-Sui speaking areas, according to the distribution map, there should be no pythons. Yet in at least two sources, names have been glossed as ‘boa’: Sui: /hui2 ɕaŋ5 B1/ (Mahidol University 2003a) and Kam: /sui212 ʔaa55/ (Mahidol University 2000). (The English gloss ‘boa’ is incorrect for although members of the family Boidae are also large constrictors, they occur only in the new world.) There is on the map an isolated population of bivittatus in eastern Sichuan, but whether it is an original part of the range, for example a northern extension of the Yunnnan range, or a human import is not clear. Because of the use of the fat and bile in Chinese traditional medicine, the animal has been widely traded (Churchman 2016, 157–159). The names given seem not to be cognate with any of the words for python found in Tai languages. Note also the recent invasive population of bivittatus in the Florida Everglades in the United States, indicating a strong propensity for adaptation to new environments for this species.
With *hnɯam A emanating from Lingnan it is worth noting that Vietnamese has not acquired this word, either from Be-Tai or from Chinese. The Vietnamese word is trăn from PMK *t( )lʌn (Muong /klan/ or /tlan/), the closest cognates found far to the south in Ha Tinh, Quang Binh and in Laos (Chamberlain 2018, 107). And although a number of Kri-Mol (Vietic) contact forms are found in Hlai, the word for python is not among them, except perhaps for Swinehoe’s nineteenth century form vang, and the Toum-Ruc (Poọng) item vang vang ‘python’ (Gérard Diffloth p.c.). It must also be the case that the Sinitic creoles of the Chinese commanderies at Jiuzhen (Thanh Hoa), Huai Huan (Nghe An), Jiude (Ha Tinh), and Rinan (Quang Binh) were established before pythons became economically important enough for the local word to have been borrowed into Early Middle Chinese. Indeed we have seen that Sek borrowed its python taxa from Kri-Mol (Vietic).
What Does Not Occur
Sometimes what is not there is just as important as what is. For example, the three main etyma for ‘pythons’ in Kra-Dai, and the Old Chinese word *prâ ‘python’ are not found in Sino-Vietnamese (*mâŋʔ may be found in literary contexts, Thiểu Chửu 2001, 586). Cognates for Chinese rán are not found in Min even though the range of bivittatus includes Fujian. Reflexes for PT *hnɯam A are not found in Kam-Sui, nor in Pu Yi, nor in Hlai, nor in Kra. And none of the words relate to Proto-Austronesian PAN *cawa.
VI Lexicon for ‘Python’ in Kra-Dai and Chinese
PAN *cawa (Wolff 2010)
Found in Philippines, Sulawesi, Old Javanese, Malay, Moken—but not Formosa
Kra
Red Gelao: ma34 ŋ̍44 ʔɗaŋ44 A ‘python, boa’ (Мазо et al. 2011)
Note: Central dialects (see below) show the B tone, the irregularity probably due to borrowing from AA.
Hlai
Proto-Hlai: *C-naaɲʔ (Cun: nɔn3, Baoting: naan3, Lauhut: naaɲ3)
(Norquest 2007) (cf Lung Ming: ŋow4 laan6 C4)
Be
Savina: niem
Zhang: num55 (A2 voiced series)
Kam-Sui
Sui: hui2 ɕaŋ5 B1 ‘boa’ (Mahidol University 2003a)
Kam: sui212 ʔaa55 ‘boa’ (Mahidol University 2000)
Sek
Gedney: ŋua4 tlɔɔ1 (older generation)
(fieldnotes) ŋua4 trɔɔ1 (younger generation)
ŋua4 trɛn1 (< Kri-Mol, eg. Ahlao: luk tǎlɛn, Chamberlain 2018, 107)
< Kri-Mol: Kri: /tălɑ̤ɑ̤/, Mlengbrou: /tăloɔ/ both mean ‘skink’
And also Mlengbrou /kra̤w/ ‘python’
Hainan
| vang | language not known (Swinehoe 1870) but curiously it seems related to Kri-Mol (Vietic), Poọng /vang vang/ (Gérard Diffloth p.c.), of the Toum-Ruc subgroup of Kri-Mol (cf Chamberlain 2018), spoken in southwestern Nghe-An Province, Vietnam. A number of Kri-Mol terms show contact with Hlai on Hainan, no doubt from the early presence of Li (Hlai) peoples on the mainland. |
Tai
PT *n̥/l̥ɯam A or *ɲ̊ɯam A
| SW: | Siamese: | lɯam A1 ‘M.r.’, laam A1 ‘P.b’ |
| Yuan: | laam A1 ‘python’ (Sanguan 1969) |
| C: | Nung: | nuom A1 (Li) |
| Tay: | lɯam A1 (initial influenced by SW dialects) | |
| WN: | num A1 (author fieldnotes) | |
| S. Zhuang: | num1, nɯm1 (Holm and Meng Yuanyao p.c.) |
| N: | Po-ai: | nɯɯm A1 (Li) |
| Yay: | nuam A1 (author fieldnotes) | |
| N. Zhuang: | tu2 nuum1 (David Holm and Meng Yuanyao p.c.) |
Central dialects only:
Lung Ming: tuu5 taaŋ5 B4 (Gedney fieldnotes)
Lung Chow, NungS: taaŋ B4 (Gedney fieldnotes)
Lei Ping: tuu2 thaaŋ5 B4 (Gedney Field notes)
NungS: tang (Savina)
S. Zhuang: tu2 taang6 B, also ‘snake’ (Holm and Meng Yuanyao p.c.)
Tày (Hoàng Văn Ma et al. 2006):
tang ‘python’
(tu) nưưm, (tu) lươm ‘earth python’
| Northern Branch: | ||
| Mashan | num1 ‘python’ ; taaŋ2 B (?) ~ ŋɯɯ ‘snake’ (Holm and Meng Yuanyao p.c.) [Mashan is situated on the boundary between Northern and Southern Zhuang.] | |
David Holm (p.c.) notes that within the Southern Zhuang area, the eastern counties such as Yongning, Shangsi, Qinzhou, Fusui and Fangcheng tend to share much of their lexicon with Northern Zhuang, whereas the western counties of Longzhou, Pingxiang, the western part of Ningming, Daxin, Dahua, and Jingxi are solidly Central Tai.
| And: | ||
| Pu Yi (Bouyei): | ŋɯ2 pan2 (Mahidol University 2001) | |
| And: | ||
| Lung ming: | ŋow4 laan6 C4 (Gedney fieldnotes) (cf. Hlai) | |
| Chinese etyma: | ||
| 蚦 | ‘python, large snake, edible’ in the Shuowen jiezi (1963, 278) [100 CE] [Probably the source for Zheng Zhang, the character 蚺 was not used until the Three Kingdoms period.] | |
| OC *njam 蚺 ‘python’ (Zheng Zhang 2013, 450) | ||
| rán ‘python’ EMC *ɲiam 蚺 (Pulleyblank 1991, 264) | ||
| naam4 (B =Tai C) ‘python’, Cantonese. Seems to have been borrowed from Tai as the tone is wrong, and therefore not directly descended from EMC. The initial agrees with Tai. | ||
| OCM | *mâŋʔ < *mlâŋʔ ‘python’ 蟒 (Schuessler 2007) |
| Amoy: bóng, báng ‘python’ (Douglas 1899) | |
| [Hainan: vang language not known (Swinehoe 1870)] |
| OCM | *prâ ‘python’ 巴 (Schuessler 2007) |
VII Conclusions
In summary, examination of the Kra-Dai words for python, enables us to infer certain patterns of Tai distribution in relation to the other families in the stock, and to confirm elements of the ordering of separations of these families from each other. It also allows us to infer that at least one of the Old Chinese terms *ɲiam was an early loan from Tai, PT *hnɯam A ‘Python bivittatu’, that probably occurred during the Han Period from Eastern Ou Yue in present day Zhekiang, in the vicinity of Wenzhou just slightly to the north of Fujian and no doubt within the range of bivittatus at the time. Be forms agree with OC with voiced onsets, and not with Tai. It is the only Kra-Dai languages to do so. As it retains the A tone, the forms are unlikely to be borrowings from Cantonese.
In terms of current geography, reflexes of PT *hnɯam A extend from the Northern and some Central dialects of Guangxi and northern Vietnam, and on through all of the Southwestern Tai dialects in Laos, Thailand, Burma, Yunnan, and Assam. For Southwestern Tai there is little variation. PSWT *hlɯam A is probably an innovation and can in most cases refer to either Python bivittatus or to Malayopython reticulatus, the exception being Siamese where the two species are differentiated by variant reflexes of the same root, /laam/ and /lɯam/. This appears to have resulted from the highly sympatric distribution of the two pythons in western and central Thailand. Elsewhere there are gaps in our data, such as Lakkia and Biao, but for the most part this is the extent of this etymon’s distribution. It has been borrowed into Cantonese, but the Min dialects to the north employ reflexes of OCM *mâŋʔ. Apart from this, the usage and geographical distribution of *mâŋʔ and of OC *prâ is unknown. It is suggested however, that the Lung Ming-Hlai correspondence, Lung Ming /laan C4/ ~ Proto-Hlai *C-naaɲʔ, may be an original Daic form that was borrowed into Old Chinese *mâŋʔ < *malâŋʔ (Schuessler 2007).
Proto-Central Tai *daaŋ reflexes are found in most Central dialects of southern Guangxi and northern Vietnam. It is cognate with Red Gelao /ʔɗaŋ44/, but the etymon appears to be an AA contact form, used often in lizard names in Khmu and Pramic, and frequently borrowed into Tai languages. The hypothesis here is that Tais received the word from Kra speakers who were the earlier inhabitants of the territory. Kra moving south from Guizhou and points north would have intermingled first with AA peoples along the Sino-Vietnamese border before heading east. There must have been some ethnic confusion between Kra and AA groups when viewed from the outside, to a degree where the term Kra became the exonym for Austroasiatic speakers in the Southwestern Tai languages, both groups residing on upland slopes and practicing swidden cultivation (for further discussion, see Chamberlain 2016, 43ff).
Thus, we may note the curious associative pattern between AA ‘skink’ and Tai ‘python’ that is manifested in AA *daaŋ ‘skink’ > CT *daaŋ ‘python’ and repeated elsewhere in AA (eg. Kri tălɑ̤ɑ̤) ‘skink’ and Sek / tlɔɔ1/ ‘python’. We might speculate, based upon Eberhard’s point mentioned above, that if the word for ‘snake’ (i.e. ‘python’) were taboo, a deceptive surrogate or avoidance term would have been needed and the skink could have been used for that purpose. It has been noted for Black Tai (Chamberlain 1977, 70) that the soul of the Calotes agamid lizard /too pɔm sakɛɛ/ is sent to heaven as a substitute for a child’s soul at the birth ceremony /sen soŋ kɯat/ to prevent the original parents from reclaiming the infant. It remains a semantic, ethnozoological, and historical linguistic puzzle.
From an examination of the folklore contained in Eberhard and the materia medica sources for Chinese as well as for Zhuang, Kam-Sui, and Gelao, it is evident that the Chinese medicinal uses of pythons developed independently and were not borrowed from Tai peoples, while the Ou Yue-Tai (NT) beliefs associated with pythons on the other hand were spiritual in nature. From their naming practices it seems the Central Branch Tais preserve clear evidence of the old taboos. And, from comparisons of the folk biological systems of the three branches, the SWT (Lo-Yue) associations with python may relate primarily to edibility. Pythons in SWT names are classed as snakes, whereas in NT they are not. CT usually avoids the name altogether and substitutes another word.
Other highly conspicuous fauna include the salt water crocodile with a similar coastal range in the north as far as Fujian. But unlike ‘python’ it has Chinese contact forms in both Fujian and Cantonese and is found throughout the Tai family with regular correspondences, PT *ŋɯak (we do not know whether reflexes occur in Be, but the Sek form is /ŋuak/ with the expected vowel correspondence). It does not occur in Kam-Sui.
Significantly, it is clear that we cannot reconstruct a root form for python of either species in Proto Kra-Dai as might be expected if Proto-Kra-Dais inhabited the territory of Lingnan in present day Guangdong and Guangxi. The range of bivittatus includes Fujian and parts of Jiangxi, but this seems to be the northern extreme of its current range. The most common root is reconstructable only in Tai. This can be interpreted to mean that the Tais moved into the core area of Lingnan only after they had separated from Kam-Sui, but before they began their migration to the west. Even Pu Yi does not have a reflex for this root. It also suggests that the origin of the Kra-Dai stock was well to the north of the range of the pythons.
The three etyma for python preserved in Tai well represent links with the other families within Kra-Dai, Hlai to the east, and Kra to the west, and even ultimately with Austroasiatic and Chinese. The northerly coastal range of the Burmese python offers support to the idea of Tai-Yue common identity expressed both linguistically and in the folklore of the snake cults described by Eberhard. I hope this brief study will lead to an appreciation of the importance of Central Tai lexicon in unraveling the complexities of the python’s position in Kra-Dai language, culture, historical geography, and external relationships, as well as of its role in demonstrating the enormous value of multidisciplinary studies.
Accepted: September 25, 2018
References
Aung Si. 2016. The Traditional Ecological Knowledge of the Solega: A Linguistic Perspective. Heidleberg: Springer.
Baxter, W. H.; and Sagart, L. 2014. Old Chinese: A New Reconstruction. New York: Oxford University Press.
Benedict, Paul. 1976. Austro-Thai. Human Relations Area Files (HRAF). New Haven: Yale University Press.
Berlin, Brent. 1992. Ethnobiological Classification: Principles of Categorization of Plants and Animals in Traditional Societies. Princeton: Princeton University Press.
―. 1972. Speculations on the Growth of Ethnobotanical Nomenclature. Language in Society 1: 51–86.
Bulmer, Ralph. 1967. Why is the Cassowary Not a Bird? A Problem of Zoological Taxonomy among the Karam of the New Guinea Highlands. Man New Series 2(1): 5–25.
Chamberlain, James R. 2018. A Kri-Mol (Vietic) Bestiary: Prolegomena to the Study of Ethnozoology in the Northern Annamites. Kyoto Working Papers on Area Studies No. 133. Kyoto University.
―. 2016. Kra-Dai and the Proto History of South China and Vietnam. Journal of the Siam Society 104: 27–77.
―. 1993. Tai-Kadai Arthropods: A Preliminary Biolinguistic Investigation. In Comparative Tai, edited by Jerold Edmondson and David Solnit, pp. 295–330. Arlington: University of Texas.
―. 1992. Biolinguistic Systematics and Marking. The Third International Symposium on Languages and Linguistics: Pan-Asiatic Linguistics vol III. Bangkok: Chulalongkorn University.
―. 1981. Proto-Tai Zoology: Serpentes. XIVth International Conference on Sino-Tibetan Languages and Linguistics. University of Florida, Gainsville.
―. 1977. An Introduction to Proto-Tai Zoology. Ph.D. Dissertation. Ann Arbor: University of Michigan.
Chi, Li. 1957. The Beginnings of Chinese Civilization. Seattle: University of Washington Press.
Churchman, Catherine. 2016. The People between the Rivers: The Rise and Fall of a Bronze Drum Culture, 200–750 CE. Lanham, Maryland: Rowman and Littlefield.
Darlington, Phillip J. 1957. Zoogeography: The Geographical Distribution of Animals. New York: Wiley.
Diffloth, Gérard. 1980. Etymological Dictionary of Mon-Khmer. Unpub. Ms.
Douglas, Carstairs. 1899. Chinese-English Dictionary of the Vernacular or Spoken Language of Amoy. London: Presbyterian Church of England.
Eberhard, Wolfram. 1968. The Local Cultures of South and East China. Leiden: Brill.
Encyclopedia Britannica. https://www.britannica.com/animal/python-snake-group. Accessed on May 17, 2018.
Fan Chengda 範成大 (Song dynasty, 1126–1193). 1984. Guihai yuheng zhi 桂海虞衡志 [Records of the supervisor and guardian of the Cassia Sea], edited by Qi Zhiping 齊治平. Nanning: Guangxi minzu chubanshe. p. 93.
Gedney, William J. 1970. The Saek Language of Nakhon Phanom Province. Journal of the Siam Society 58 (pt.1): 67–87.
Groot, T. V. M.; Bruins, E.; and Breeuwer, J. A. J. 2003. Molecular Genetic Evidence for Parthenogenesis in the Burmese Python, Python molurus bivittatus. Heredity 90: 130–135.
Hansell, Mark. 1988. The Relation of Be to Tai: Evidence from Tones and Initials. In Comparative Kadai: Linguistic Studies beyond Tai, edited by J. Edmondson and D. Solnit, pp. 239–288. Arlington: University of Texas and Summer Institute of Linguistics.
Hoàng Văn Ma; Lục Văn Pảo; and Hoàng Chí. 2006. Từ điển Tày-Nùng-Việt [Tay-Nung-Vietnamese Dictionary]. Hà Nội: Nhà xuất bản Từ điển bách khoain Ba Đình.
Holm, David. 2017. Vernacular Character Scripts of the Zhuang, Tày, and Nùng: Preliminary Comparisons. Paper presented at the International Conference on Sino-Tibetan Languages and Linguistics (ICSTLL), Chinese Academy of Social Sciences.
―. 2015. A Layer of Old Chinese Readings in the Traditional Zhuang Script. Bulletin of the Museum of Far Eastern Antiquities 79/80: 169–220.
Huang Hanru 黄汉儒, ed. 2001. Zhongguo Zhuang Yixue 中国壮医学 [Medicine of China’s Zhuang]. Nanning: Guangxi minzu chubanshe.
Huang Huaixin 黄懷信 et al. 2007. Yi Zhou shu huijiao jizhu 逸周書彙校集注 [The Left-over Zhou Documents: Assembled recensions and collected notes], revised edition. Shanghai: Shanghai guji chubanshe.
Li, Fang Kuei. 1977. A Handbook of Comparative Tai. Honolulu: University of Hawai’i Press.
Liang Qicheng 梁启成; and Zhong Ming 钟鸣. 2005. Zhongguo Zhuang Yaoxue 中国壮药学 [Pharmacology of China’s Zhuang]. Nanning: Guangxi minzu chubanshe.
Lin Luhe 林呂何, ed. 1991. Guangxi yaoyong dongwu 廣西藥用動物 [Guangxi medicinal animals]. Nanning: Guangxi kexue jishu chubanshe.
Мазо O. M. Mazo O. M.; Нгуен В. Л. Nguen V. L.; Нгуен Х. Х. Nguen H. H.; and Самарина И. В. Samarina I. V. 2011. Jazyki gèlao: Materialy k sopostavitel’nomu slovarju kadajskih jazykov Языки гэлао: материалы к сопоставительному словарю кадайских языков [Gelao Languages: Materials for a comparative dictionary of the Kadai languages]. Moscow: Academia.
Mahidol University. 2003a. Sui, Chinese Thai, English Dictionary. Bangkok: Mahidol University, Institute of Language and Culture for Rural Development.
―. 2003b. Hlai (Li), Chinese, Thai, English Dictionary. Bangkok: Mahidol University, Institute of Language and Culture for Rural Development.
―. 2001. Bouyei, Chinese, English, Thai Dictionary. Bangkok: Mahidol University, Institute of Language and Culture for Rural Development.
―. 2000. Kam, Chinese, Thai, English Dictionary. Bangkok: Mahidol University, Institute of Language and Culture for Rural Development.
McKinnon, J.; and McKinnon, K. 1974. Animals of Asia: the Ecology for the Oriental Region. New York: Holt, Reinhart and Winston.
Miyake, Marc. 2018. http://www.amritas.com/180512.htm#05112359. 18.5.11.23:59: Chu And Kra-Dai (Part 2). Accessed on June 2, 2018.
Müller, Paul. 1974. Aspects of Zoogeography. The Hague: Junk.
Norquest, Peter K. 2007. A Phonological Reconstruction of Proto-Hlai. Ph.D. Dissertation, University of Arizona.
Ostapirat, Weera. 2018. Faunal Terms in Kra-Dai and Austro-Tai. Paper presented at the Special Workshop on Faunal Lexicons in Mainland Southeast Asia. Kobe, Japan.
―. 2013. The Rime System of Proto-Tai. Bulletin of Chinese Linguistics 7(1): 189–227.
―. 2008. The Hlai Language. In The Tai-Kadai Languages, edited by Anthony V. N. Diller and Jerold A. Edmondso, pp. 623–652. New York and London: Routledge.
―. 2004. Proto-Hlai Sound System and lexicons. In Studies on Sino-Tibetan Languages: Papers in Honor of Professor Hwang-cherng Gong on His Seventieth Birthday, edited by Yin-chin Lin, Fang-min Hsu, Chǔn-chih Lee, Jackson T.-S. Sun, Hsiu-fang Yang, and Dah-an Ho, pp. 121–175. Taipei: Institute of Linguistics. Academia Sinica.
Pan Lutai 潘炉台 et al., eds. 2003. Buyizu yiyao 布依族医药 [Buyi family medicine]. Guiyang: Guizhou minzu chubanshe.
Pulleyblank, Edwin G. 1991. Lexicon of Reconstructed Pronunciation in Early Middle Chinese, Late Middle Chinese, and Early Mandarin. Vancouver: University of British Columbia Press.
―. 1984. Middle Chinese: A Study in Historical Phonology. Vancouver: University of British Columbia Press.
Qin Xunyun 覃迅云; and Li Tong 李彤, eds. 2001. Zhongguo Yao yixue 中国瑶医学 [Medicine of China’s Yao]. Beijing: Minzu chubanshe.
Sanguan Chootsukarat สงวน โชติสุขรัตน์. 1969. Thay Yuan-Khon Meuang ไทยวน-คนเมือง [Thai Yuan-Meuang people]. Chonburi: Bang Sare Printers.
Savina, F. M. 1965. Vocabulaire Bê [Be vocabulary], edited by A. Haudricourt. Paris: EFEO.
―. 1931. Lexique Đai-Français [Dai-French lexison]. Hanoi.
―. 1924. Dictionnaire Etymologique Français-Nung-Chinois [French-Nung-Chinese etymological dictionary]. Hong Kong.
―. 1910. Dictionnaire Tày-Annamite-Français [Tay-Vienamese-French dictionary]. Hanoi: Imp. d’Extrême Orient.
Schafer, Edward. 1967. The Vermilion Bird. Berkeley: University of California Press.
Schuessler, Axel. 2007. ABC Etymological Dictionary of Old Chinese. Honolulu: University of Hawai‘i Press.
Shuowen jiezi 13A:16a. 1963. Beijing: Zhonghua shuju edition.
Solnit, David B. 1988. The Position of Lakkia within Kadai. In Comparative Kadai: Linguistic Studies Beyond Tai, edited by J. Edmondson and D. Solnit, pp. 219–238. Arlington: University of Texas and Summer Institute of Linguistics.
Stuart, B.; Nguyen, T. Q.; Thy, N.; Grismer, L.; Chan-Ard, T.; Iskandar, D.; Golynsky, E.; and Lau, M. W. N. 2012. Python bivittatus. The IUCN Red List of Threatened Species 2012: e.T193451A2237271. http://dx.doi.org/10.2305/IUCN.UK.2012-1.RLTS.T193451A2237271.en.
Swinehoe, R. 1870. List of Reptiles and Batrachians Collected in the Island of Hainan (China), with Notes. Proc. Zool. Soc. London: 239–241.
Thiều Chửu, ed. 2001. Hán-Việt Tự Điển [Sino-Vietnamese dictionary], TP. Hồ Chí Minh: Nhà xuất bản TP.
Wang Bocan 王柏灿, ed. 2006. Lidai Zhuangzu yiyao shiliao huicui 历代壮族医药史料荟萃 [Digest of historical materials on Zhuang medicine through the ages]. Nanning: Guangxi minzu chubanshe.
Wang Hou’an 王厚安, ed. 1997. Shuizu yiyao 水族医药 [Shui Nationality’s Medicine]. Guiyang: Guizhou minzu chubanshe.
Wolff, John U. 2010. Proto-Austronesian Phonology with Glossary. Ithaca: Cornell University, Southeast Asia Program Publications.
Zhang Yuangsheng 张元生; Ma Jialing 马嘉陵; Wen Mingying 温明英; and Wei Xinglang 和魏兴朗. 1985. Hainan Lingao hua 海南岭澳华 [The language of Lingao, Hainan]. Nanning: Guangxi Nationalities Press.
Zhao Junhua 赵俊华 et al., eds. 2003. Gelaozu yiyao 仡佬族医药 [Gelao ancestral medicine]. Guiyang: Guizhou minzu chubanshe.
Zheng Zhang, Shangfang 鄭張尚芳. 2013. Shanggu Yinxi 上古音系 [Phonetic system of Archaic Chinese]. Shanghai: Jiaoyu chubanshe.
Zhu Hua 朱华, ed. 2003. Zhongguo Zhuang yaozhi 中国壮药志 [Pharmacopaedia of China’s Zhuang], vol. 1. Nanning: Guangxi minzu chubanshe.
![]()


{kind=link}